To use all functions of this page, please activate cookies in your browser.
My watch list
my.chemeurope.com
my.chemeurope.com
With an accout for my.chemeurope.com you can always see everything at a glance – and you can configure your own website and individual newsletter.
- My watch list
- My saved searches
- My saved topics
- My newsletter
Peroxisome
Peroxisomes are ubiquitous organelles in eukaryotes that participate in the metabolism of fatty acids and other metabolites. Peroxisomes have enzymes that rid the cell of toxic peroxides. They have a single lipid bilayer membrane that separates their contents from the cytosol (the internal fluid of the cell) and contain membrane proteins critical for various functions, such as importing proteins into the organelles and aiding in proliferation. Like lysosomes, peroxisomes are part of the secretory pathway of a cell, but they are much more dynamic and can replicate by enlarging and then dividing. Peroxisomes were identified as cellular organelles by the Belgian cytologist Christian de Duve in 1967 after they had been first described in a Swedish PhD thesis a decade earlier. Additional recommended knowledge
Occurrence and evolutionPeroxisomes are found in virtually all eukaryotic cells and stem cells. Peroxisomes contain enzymes for certain oxidative reactions, like the beta-oxidation of very-long-chain fatty acids. Prokaryotes lack peroxisomes. The enzymatic content of peroxisomes varies across species, but the presence of certain proteins common to many species has been used to suggest an endosymbiotic origin; that is, peroxisomes evolved from bacteria that invaded larger cells as parasites, and very gradually evolved a symbiotic relationship.[1] However, this view has been challenged by recent discoveries. For example, peroxisome-less mutants can restore peroxisomes upon introduction of the wild-type gene, and peroxisomes have been observed to be formed from the endoplasmic reticulum.[2] An evolutionary analysis of the peroxisomal proteome found homologies between the peroxisomal import machinery and the ERAD pathway in the endoplasmic reticulum, along with a number of metabolic enzymes that were likely recruited from the mitochondria.[3] These results indicate that the peroxisome does not have an endosymbiotic origin; instead, it likely originates from the ER, and its proteins were recruited from pools existing within the primitive eukaryote, as quoted in the science textbook Biozone. FunctionPeroxisomes contain oxidative enzymes, such as catalase, D-amino acid oxidase, and uric acid oxidase.[4] Certain enzymes within the peroxisome, by using molecular oxygen, remove hydrogen atoms from specific organic substrates (labeled as R), in an oxidative reaction, producing hydrogen peroxide (H2O2, itself toxic): catalase, another enzyme in the peroxisome, in turn uses this H2O2 to oxidize other substrates, including phenols, formic acid, formaldehyde, and alcohol, by means of the peroxidation reaction:
This reaction is important in liver and kidney cells, where the peroxisomes detoxify various toxic substances that enter the blood. About 25% of the ethanol we drink is oxidized to acetaldehyde in this way. In addition, when excess H2O2 accumulates in the cell, catalase converts it to H2O through this reaction: A major function of the peroxisome is the breakdown of fatty acid molecules, in a process called beta-oxidation. In this process, the fatty acids are broken down two carbons at a time, converted to Acetyl-CoA, which is then transported back to the cytosol for further use. In animal cells, beta-oxidation can also occur in the mitochondria. In yeast and plant cells, this process is exclusive for the peroxisome. The first reactions in the formation of plasmalogen in animal cells also occurs in peroxisomes. Plasmalogen is the most abundant phospholipid in myelin. Deficiency of plasmalogens causes profound abnormalities in the myelination of nerve cells, which is one of the reasons that many peroxisomal disorders lead to neurological disease. Peroxisomes also play a role in the production of bile acids and proteins. Protein importProteins are selectively imported into peroxisomes. Since the organelles contain no DNA or ribosomes and thus have no means of producing proteins, all of their proteins must be imported across the membrane. It is believed that proteins do not transit through the endoplasmic reticulum to get to the peroxisome. A specific protein signal (PTS or peroxisomal targeting signal) of three amino acids at the C-terminus of many peroxisomal proteins signals the membrane of the peroxisome to import them into the organelle. Other peroxisomal proteins contain a signal at the N-terminus. There are at least 32 known peroxisomal proteins, called peroxins, which participate in the process of importing proteins by means of ATP hydrolysis. Proteins do not have to unfold to be imported into the peroxisome. The protein receptors, the peroxins Pex5 and Pex7, accompany their cargoes (containing a PTS1 or a PTS2, respectively) all the way into the peroxisome where they release the cargo and then return to the cytosol - a step named recycling. Overall, the import cycle is referred to as the extended shuttle mechanism. Evidence now indicates that ATP hydrolysis is required for the recycling of receptors to the cytosol. Also, ubiquitination appears to be crucial for the export of PEX5 from the peroxisome, to the cytosol. Little is known about the import of PEX7, although it has helper proteins that have been shown to be ubiquitinated. DeficienciesPeroxisomal disorders are a class of condtions that lead to disorders of lipid metabolism. One well-known example is Zellweger syndrome. Peroxisomes matrix proteins are synthesized on free ribosomes in the cytosol and that these proteins are imported posttranslationally in pre-existing peroxisomes. References
This article contains material from the Science Primer published by the NCBI, which, as a U.S. government publication, is in the public domain.
|
|||
| This article is licensed under the GNU Free Documentation License. It uses material from the Wikipedia article "Peroxisome". A list of authors is available in Wikipedia. |



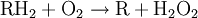
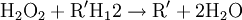 , thus eliminating the poisonous hydrogen peroxide in the process.
, thus eliminating the poisonous hydrogen peroxide in the process.