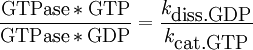To use all functions of this page, please activate cookies in your browser.
My watch list
my.chemeurope.com
my.chemeurope.com
With an accout for my.chemeurope.com you can always see everything at a glance – and you can configure your own website and individual newsletter.
- My watch list
- My saved searches
- My saved topics
- My newsletter
GTPase
They help GTP binding proteins hydrolyse GTP and be converted to their ground state. Product highlight
FunctionsGTPases play an important role in:
MechanismThe hydrolysis of the γ phosphate of GTP into guanosine diphosphate (GDP) and Pi,inorganic phosphate, occurs by the SN2 mechanism (see nucleophilic substitution) via a pentavalent intermediate state and is dependent on the magnesium ion Mg2+. Major MotifsIn most GTPases, the specificity for the base guanine is imparted by the base-recognition motif, which has the consensus sequence [N/T]KXD. [1] Regulatory GTPasesRegulatory GTPases, also called the GTPase superfamily, are GTPases used for regulation of other biochemical processes. Most prominent among the regulatory GTPases are the G proteins. GTP switchAll regulatory GTPases have a common mechanism that enables them to switch a signal transduction chain on and off. Throwing the switch is performed by the unidirectional change of the GTPase from the active, GTP-bound form to the inactive, GDP-bound form by hydrolysis of the GTP through intrinsic GTPase-activity, effectively switching the GTPase off. This reaction is initiated by GTPase-activating proteins (GAPs), coming from another signal transduction pathway. It can be reverted (switching the GTPase on again) by Guanine nucleotide exchange factors (GEFs), which cause the GDP to dissociate from the GTPase, leading to its association with a new GTP. This closes the cycle to the active state of the GTPase; the irreversible hydrolysis of the GTP to GDP forces the cycle to run only in one direction. Only the active state of the GTPase can transduce a signal to a reaction chain. Switch regulationThe efficiency of the signal transduction via a GTPase depends on the ratio of active to inactive GTPase. That equals:
with kdiss.GDP being the dissociation constant of GDP, and kcat.GTP the hydrolysis constant of GTP for the specific GTPase. Both constants can be modified by special regulatory proteins.
Heterotrimeric G proteinsThese G proteins are made from three subunits, with the G domain located on the largest one (the α unit); together with the two smaller subunits (β and γ units), they form a tightly associated protein complex. α and γ unit are associated with the membrane by lipid anchors. Heterotrimeric G proteins act as the specific reaction partners of G protein-coupled receptors. The GTPase is normally inactive. Upon receptor activation, the intracellular receptor domain activates the GTPase, which in turn activates other molecules of the signal transduction chain, either via the α unit or the βγ complex. Among the target molecules of the active GTPase are adenylate cyclase and ion channels. The heterotrimeric G proteins can be classified by sequence homology of the α unit into four families:
By combination of different α, β and γ subunits, a great variety (>1000) G proteins can be produced. GDP is not needed for GTP. Activation cycle of heterotrimeric G proteinsIn the basic state, the Gα-GDP-Gβγ complex and the receptor that can activate it are separately associated with the membrane. On receptor activation, the receptor becomes highly affine for the G protein - GDP complex. On binding with the complex, GDP dissociates from the complex; the receptor works as a GEF - GDP-GTP Exchange Factor; the free complex has a high affinity for GTP. Upon GTP binding, both Gα-GTP and Gβγ separate from both the receptor and from each other. Depending on the lifetime of the active state of the receptor, it can activate more G proteins this way. The Ras GTPase superfamilyThese are small monomeric proteins homologous to Ras. They are also called small GTPases. Small GTPases have a molecular weight of about 21 kilo-Dalton and generally serve as molecular switches for a variety of cellular signaling events. According to their primary amino acid sequences and biochemical properties, the Ras superfamily is further divided into five subfamilies: Ras, Rho, Rab, Arf and Ran. Translation factor familyThese GTPases play an important role in initiation, elongation and termination of protein biosynthesis. Translocation factorsSee signal recognition particle (SRP). Large GTPasesSee dynamin as a prototype for large GTPases. See also
References
Categories: Signal transduction | EC 3.6.5 |
|||||||||||||||||||
| This article is licensed under the GNU Free Documentation License. It uses material from the Wikipedia article "GTPase". A list of authors is available in Wikipedia. | |||||||||||||||||||