To use all functions of this page, please activate cookies in your browser.
My watch list
my.chemeurope.com
my.chemeurope.com
With an accout for my.chemeurope.com you can always see everything at a glance – and you can configure your own website and individual newsletter.
- My watch list
- My saved searches
- My saved topics
- My newsletter
Ribonuclease A
Ribonuclease A (RNase A) is an endonuclease that cleaves single-stranded RNA. Bovine pancreatic RNase A is one of the classic model systems of protein science.
Product highlight
HistoryThe importance of bovine pancreatic RNase A was secured when the Armour & Co. (of hot dog fame) purified a kilogram of it, and gave 10 mg samples away free to any interested scientists. The ability to have a single lot of purified enzyme instantly made RNase the model system for protein studies. RNase A was the model protein used to work out many spectroscopic methods for assaying protein structure, including absorbance, circular dichroism/optical rotary dispersion, Raman, EPR and NMR spectroscopy. RNase A was also the first model protein for the development of several chemical structural methods, such as limited proteolysis of disordered segments, chemical modification of exposed side chains, and antigenic recognition. RNase A was the third protein to have its structure solved, in 1967.[1] Studies of the oxidative folding of RNase A led Chris Anfinsen to enunciate the thermodynamic hypothesis of protein folding, which states that the folded form of a protein represents the minimum of its free energy. RNase A was the first protein for showing the effects of non-native isomers of X-Pro peptide bonds in protein folding. RNase A was the first protein to be studied by multiple sequence alignment and by comparing the properties of evolutionarily related proteins. Structure and Properties
RNase A has four disulfide bonds in its native state: Cys26-Cys84, Cys58-110, Cys40-95 and Cys65-72. The first two (26-84 and 58-110) are essential for conformational folding; each joins an alpha helix of the first layer to a beta sheet of the second layer, forming a small hydrophobic core in its vicinity. The latter two disulfide bonds (40-95 and 65-72) are less essential for folding; either one can be reduced (but not both) without affecting the native structure under physiological conditions. These disulfide bonds connect loop segments and are relatively exposed to solvent. Interestingly, the 65-72 disulfide bond has an extraordinarily high propensity to form, significantly more than would be expected from its loop entropy, both as a peptide and in the full-length protein. This suggests that the 61-74 β-hairpin has a high propensity to fold conformationally.
The N-terminal α-helix of RNase A (residues 3-13) is connected to the rest of RNase A by a flexible linker (residues 16-23). As shown by F. M. Richards, this linker may be cleaved by subtilisin between residues 20 and 21 without causing the N-terminal helix to dissociate from the rest of RNase A. The peptide-protein complex is called RNase S, the peptide (residues 1-20) is called the S-peptide and the remainder (residues 21-124) is called the S-protein. The dissociation constant of the S-peptide for the S-protein is roughly 30 pM; this tight binding can be exploited for protein purification by attaching the S-peptide to the protein of interest and passing a mixture over an affinity column with bound S-protein. [A smaller C-peptide (residues 1-13) also works.] The RNase S model system has also been used for studying protein folding by coupling folding and association. The S-peptide was the first peptide from a native protein shown to have (flickering) secondary structure in isolation (by Klee and Brown in 1967).
Enzymatic MechanismThe positive charges of RNase A lie mainly in a deep cleft between two lobes. The RNA substrate lies in this cleft and is cleaved by two catalytic histidines, His12 and His119, via a Anti-cancer effectsRNase A, and to a greater extent its oligomers and some homologs (such as onconase from frogs), have cytotoxic and cytostatic effects, particularly on cancer cells. This has led to the development of onconase as a cancer therapeutic, particularly for external use against skin cancers. As with many protein drugs, the internal use of non-human ribonucleases such as onconase is limited by the patient's immune response. Other biological effectsRibonuclease is also related to angiogenin, which is involved in blood vessel development. See also
References
Categories: Ribonucleases | EC 3.1.27 |
|||||||||||||||||||||||
| This article is licensed under the GNU Free Documentation License. It uses material from the Wikipedia article "Ribonuclease_A". A list of authors is available in Wikipedia. | |||||||||||||||||||||||



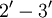 cyclic phosphate intermediate that is stabilized by nearby lysines such as Lys7, Lys41 and Lys66.
cyclic phosphate intermediate that is stabilized by nearby lysines such as Lys7, Lys41 and Lys66.


